
Effects of Night Illumination on Behavior, Body Mass and Learning in Male Zebra Finches
by
 , ,
, ,
Abhilash Prabhat
1 ,
,
Mayank Kumar
1,†,
Ashwani Kumar
1,†,
Vinod Kumar
2 and
Sanjay K. Bhardwaj
1,* 1
Department of Zoology, Chaudhary Charan Singh University, Ramgarhi, Meerut 250001, Uttar Pradesh, India
2
Department of Zoology, University of Delhi, Delhi 110007, India
*
Author to whom correspondence should be addressed.
†
Equal contribution.
Birds 2021, 2(4), 381-394; https://doi.org/10.3390/birds2040028
Submission received: 7 October 2021
/
Revised: 9 November 2021
/
Accepted: 10 November 2021
/
Published: 12 November 2021
(This article belongs to the Special Issue Feature Papers of Birds 2021)
Abstract
:Simple Summary
A temporal disrupted environment has the ability to enable activities that allow animals (including humans) to eat, forage, and sleep throughout the day. It can lead to circadian disruption and altered health conditions. We exposed male zebra finches to dim light at night (dLAN) for six weeks and assessed their daily behavioral patterns, sleep behavior, physiology, object exploration, and learning and memory. Dim light at night altered 24-h behavioral rhythms and negatively affected sleep behavior, color learning, and memory performance. Specifically, the feeding behavior became arrhythmic and birds gained fat, leading to increase in body mass. We suggest that this could be the result of change in rhythmicity and allocation of energy expenditure throughout the day. Alteration in rhythm and sleep loss affected overall mood and motivation to interact with novel objects (similar to neophobia) and increased non-performance in colour learning and memory tasks under dLAN. Insights from these results may be useful in understanding the impact of nighttime illumination, such as in an overly lit urban habitat, on daily behavior, physiology, and advanced brain functions in animals.
Abstract
An increase in artificial night lighting has blurred the boundaries of day and night and transformed the natural day-night environment with alteration in the temporal niche of the animals. Male zebra finches were exposed to a dim light at night (dLAN) protocol (Light: dLAN, 12L = 200 lux: 12dLAN = 5 lux) with controls on darkness at night (Light: dark, 12L = 200 lux: 12D = 0 lux) for six weeks. We assayed sleep-wake, daily behaviors, mood, and cognition, as well as changes in physiological parameters. Dim light at night increased sleep frequency, delayed sleep onset, advanced awakening latency, and caused a reduction in total sleep duration. dLAN birds did not associate (physical association) with novel object and birds spent significantly lesser time on perch with novel object as compared to LD. In colour learning task, night illuminated birds took more time to learn and made more error, compared to LD. dLAN significantly altered the 24-h daily behavioral rhythm (amplitude and acrophase) of feeding, drinking, preening, and perch-hopping behavior. In particular, birds extended their feeding hours in the nighttime under dLAN, with no difference in total food intake. Birds under dLAN increased fattening and hence significantly increased body mass. Our results show that dim light at night altered feeding rhythm, caused decrease in sleep behavior, and negatively affected learning and memory performance in male zebra finches.
1. Introduction
Like all animals, birds show adaptation to the light environment and show distinct physiological and behavioral differences between day and night. The presence and absence of light between day and nighttime, respectively, has a major role in the functioning of endogenous clock and its output behavior [1,2,3,4,5]. In the last decade there has been increasing emphasis on the investigation of the impact of light at night (LAN) which causes a mismatch of internal (endogenous clocks) and external (environmental cue) timing on the biology of animals and plants. It has reduced the darkness of night, and the differences between day-night periods are blurred [6]. This has affected the synchronization of clock with clock-controlled functions such as daily circadian behaviors, hormone secretions, metabolism, and cognition across the animal kingdom [7,8,9,10]. Emerging evidence suggests an association with LAN-induced circadian disruptions, with increasing incidences of loss of sleep behavior, mood disorder, changes in body physiology, and an adverse effect on reproduction in both diurnal and nocturnal vertebrates [11,12,13,14].
Recent examples in diurnal songbirds have shown the effects of light at night at the behavioral, physiological and molecular levels. Specifically, increased light at night disrupted daily rhythm with loss of nocturnal peak in melatonin secretion in the Indian weaver bird (Ploceus philippinus; [15]), great tits (Parus major; [8,9,10,11,12,13,14,16]), Indian house crows (Corvus splendens; [14]) and female zebra finches (Taeniopygia guttata; [17,18]). Bright light at night (constant light) and removal of pineal gland exposure decayed daily rhythms in singing and activity behaviors in zebra finches [13,19,20,21] and alters the perception of day length and affects the underlying physiology in Indian Weaver bird [22]. The light at night induces “eating at a wrong time” in both mammals and birds. Evidence shows that LAN alters food intake timing and metabolic rhythms and consequently body mass gain and obesity in rats [7,8,23] and zebra finches [17]. Acute and chronic dim light at night (dLAN) exposure led to the obesity and body mass gain with increased fat score without alteration in the total daily caloric intake in rats and female zebra finches [8,17].
Light pollution has greatly contributed to the sleep debt in free-living and captive birds [14,16,18,24]. No-night environment and dim light at night caused negative effects on advance brain functions such as acquisition of learning and memory and novel object exploration, and hence cognition in zebra finches and Indian house crows [13,14]. Zebra finches increased the number of trials in cognitive tasks and negatively correlated with the total song bouts and unique syllables [13]. In Indian house crows, constant light induced circadian rhythm disruption and impairment in spatial and pattern learning [10]. These cognitive behaviors are hippocampus-dependent and studies on higher cognitive function implicates brain health of an individual [14,25]. Few studies have shown the chronic effect of dim light at night (as constant light is not relevant at the ecological level; Jha and Kumar, 2017 [13]) on sleep, mood and cognitive performance in zebra finches.
Zebra finches are diurnal and highly social avian species that can breed well in the laboratory conditions and show all behavioral responses in captivity such as feeding, perch hopping and reproductive behavior [13,26,27]. It is an excellent model to study behavior and physiology and widely used for understanding highly focused and mechanistic questions [28]. We designed the present study similar to Batra et al., (2019) [17] in which only females are subjected to 5 lux of dim light at night. We used similar protocol on male zebra finches instead and replicated the effects on daily behavior, physiology and sleep. We also investigated effects of dLAN on novel object exploration, colour learning and memory task. We expect that daily behavioral rhythm of male zebra finches will be disturbed by light at night and changes in feeding behavior will lead to the fattening and increase in body mass. We also speculate that dim night illumination of six weeks will impair the learning and memory in these birds as shown in constant light and dim light at night condition in zebra finches and Indian house crows [10,13].
2. Materials and Methods
2.1. Animal Husbandry and Experimental Protocol
The study was approved by the Institutional Animal Ethics Committee of the Department of Zoology, Chaudhary Charan Singh University, Uttar Pradesh, India. We used adult zebra finches that were born and raised in our indoor breeding colony under 12 h photoperiod (12 h light: 12 h darkness, 12L:12D; L ~ 200 lux, D < 0.1 lux) and 22 ± 2 °C temperature. The compact fluorescent lamps (5W, 220–240 V: Crompton, India) provided light illumination and a Mueller automatic timer (model SC 88) controlled the timing of light and dark periods. The lamp produced white light which encompasses all the visible colours (cool daylight with colour temperature 6500 K) and used in almost all studies in the laboratory. All birds received foxtail millet (Setaria italica) seeds as main food, hard-boiled eggs mashed with crushed eggshells, fresh vegetables and fruits and grit as daily diet supplement [13].
This study was carried out in the month January and February with controlled lighting; light:dark (12L:12D) and temperature (22 ± 2 °C) to remove the effects of season, if any. Adult male zebra finches (n = 16) were singly housed in aluminium mesh cage (46 cm × 40 cm × 46 cm) placed inside a light and soundproof box (81 cm × 68 cm × 66 cm) in similar light and food conditions. Similar protocols have been used in many publications for songbirds in which confinement to a long duration did not show any deleterious effect on health of birds [14,17,25]. After a week of acclimatization to the new cage environment, randomly half of the birds were provided with dim light at night (12 h light: 12 h dim light at night, LAN, 12L:12dLAN; L = 200 ± 10 lux, dLAN = 5 ± 1 lux) of similar white light (cool daylight with colour temperature 6500 K) and other half on similar lighting condition (12L:12D; L = 200 ± 10 lux, D < 0.1 lux) i.e., darkness at night for six weeks (Figure 1; Experimental Protocol). We measured night light intensity near university area and used Philips (India, 220–240 V) to provide LAN. The light intensity was measured using Macam Q203 Quantum (Scotland, UK) radiometer. Food and water were available ad libitum for both groups [14,17,18].
2.2. Measurement of Body Mass, Food Intake, Seed Storage in Crop and Fat Score
We measured body mass, fat score, and food intake at the start and end of the experiment (Figure 1). Birds were weighed on a table top balance to an accuracy of 0.1 g in the morning. Food intake was measured as per Prabhat et al., (2020) [27]. After six weeks, we recorded body subcutaneous fat depots over tracheal pit (T) and abdomen (A) regions and assessed and scored in a scale of 0–4, where 0 = no visible fat (dark red); 1 = T, wide wedge of fat; A, trace of fat (light red); 2 = T, completely covered with fat but deeply concave; A, slips of fat (light yellow); 3 = T, moderate fat reserves but concave; A, slightly bulging pad (light yellow); 4 = T, heavy fat deposits up to clavicles; A, clearly bulging fat pad (yellow). We also monitored changes in crop seed content as a marker to monitor stored feed, and we counted seeds visible on the surface of both left and right crops at the start and end of the experiment at lights on [17,27].
2.3. Assay of Daily Behavior
We assayed daily feeding, drinking, perch hopping, and preening behavior in both the light conditions (n = 8/light condition). We videographed zebra finch behavior over two consecutive days at the end of week five using night vision full HD camera (Figure 1). We mounted video camera at the center top of the cage, so that whole cage is visible which enables us to observe postures and activities of bird. All videographs were scanned manually by a researcher unaware of the experimental protocol to remove experimenter’s bias. We calculated frequency (how many times) and duration (how long) of feeding (when bird actually ate) and drinking behavior, perch-hopping (bird’s directed activity from one perch to the other) and preening behavior (self-grooming) for 24 h over two days and averaged it. We arranged all behavioral data in half an hour bin and used it for rhythm analysis. We plotted non-linear cosinor regression line of each behavior for both light conditions to look at the effect of light at night on rhythmic patterns. For further quantification, we totaled every day’s behavioral data for each individual bird and presented in mean (±SE) for both LD and dLAN and compared day vs. night values.
2.4. Behavioral Assay of Sleep
At the end of the experiment, we monitored the posture birds by night vision IR HD camera and recorded it as sleep or awake state [14,18,29,30,31]. Later, these videos were analyzed on the basis of postures (i.e., front and back sleep) [14,18], where front sleep state was bird with closed eyes and head in front whereas back sleep was a state with head tucked back on scapula [14,18]. We assayed sleep latency (interval from light off to sleep onset), awakening latency (interval from sleep offset to light on), bout durations and total sleep via video recordings. The scores obtained from videographs over two days were averaged, and from this mean (±s.e.) bout duration of sleep behavior was calculated. A sum of all sleep bout gave total sleep duration at night in both the light conditions.
2.5. Novel Object Exploration (NOE) Test
To investigate the effect of dim light at night on mood, we measured the exploratory response of male Zebra Finches to a novel object, since it is a measure of mood state and loss of interest in performing brain-dependent functions [14,32]. The exploratory response was tested in the same cage in which these birds were singly housed caged to minimize the novelty of new cage environment between 4 to 8 h after lights on. Before testing, clean blotting sheets were placed on the floor of cages along with water and food. After 15 min of acclimation, a nonfood novel object (NO, a pink triangular prism shaped eraser, l = 29 mm, b = 12.5 mm, h = 11.8 mm) was fixed on a randomly chosen perch, in a manner that it was placed equidistance from centrally placed food and water cups. The exploratory response towards this novel object was quantified for 10 min, by analyzing the videographed data. We analyzed approach score towards NO, time spent on perch with NO, number of perch hops, and time taken to closely associate with novel object. We quantified proximity to the novel object as approach score on a scale of 0–5 [13,32] where 0: bird on the floor at furthest distance from the NO; 1: bird on floor in the center of two perches and equidistant from the NO; 2: bird on floor but towards perch with NO; 3: bird on the perch without NO; 4: bird on the perch with NO; 5: bird on perch with novel object and closely associating with it (touching or pecking) (Video S1).
2.6. Colour Learning and Memory Retrieval
We also tested effect of dim light at night on cognitive abilities of these male zebra finches. Both the groups were tested for colour learning and memory retrieval task. This task required a bird to associate between rewarded cup and lid colour as well as to ignore the unrewarded lid during memory retrieval test. To remove the spatial bias, we randomly changed the position of rewarded cup in each consecutive trial. A white Bakelite sheet (78.5 mm × 152.5 mm) was used as base of test apparatus, on which six plastic food wells (20 mm wide × 14 mm deep) were pasted in two rows of three food wells. Lids used in this task was made of coloured sheet (0.1 g) of diameter 26 mm with a metal plate (0.4 g) was attached to lids; each lid thus weighed at 0.5 ± 0.1 g. We used orange and navy blue-colored lids (one rewarded and 5 unrewarded colours). To gain the food reward (fox tail millet) all the bird needed to do was flip the lid off the food cup of apparatus either by pecking or by their feet (Videos S2 and S3). The task was similar to Boogert et al., (2008) [33]; Jha and kumar (2017) [13] with appropriate modification.
At the end of the experiment (Figure 1), birds received food in the task apparatus instead of the regular food cup to reduce neophobia, if any, and to familiarize test subjects with the apparatus. On the next day, food was removed from the cages before lights off, to give a food deprivation period of 15 h. Finally, on day three these birds were tested for colour learning task. We standardized the trial period of 10 min in our pilot experiment, so the test required was to find the rewarded coloured cup within 10 min of introduction of apparatus in the cage. A successful bird was allowed to eat for only 10 s, whereas an unsuccessful individual was given free food access. We performed and filmed a maximum of 15 trials continuously for each bird during training period, if required. A successful bird was one that performed the task without an error five times in a row in learning period [13] (Supplementary video). After the last trial, the successful bird was again given a food deprivation for 1 h to test 1 h retention interval (RI) memory. In RI, birds were tested for only five trials of maximum 10 min and after those birds were given food ad libitum for the remainder of the day and again food deprived for 15 h to test 24 h memory retention on the next day. Birds who did not participate received food ad libitum for the remainder of the day. The successful completion of first step meant progress to the next one, however failure to do so reverted the individual to the previous step. Thus, birds learnt the correct rewarded well by trial and error. We controlled for inadvertent visual and olfactory cues by preparing food wells out of birds’ sight each time. Videos were recorded for all behavioral tests and analyzed manually by a researcher unaware of the experimental conditions to remove any experimenter’s bias.
2.7. Statistics
All statistical analyses were performed using GraphPad prism software, version 6.0 (La Jolla, CA, USA). Unpaired parametric Student’s t-tests compared the physiological parameters (changes in body mass, food intake and crop seed content), quantification of daily behaviors (feeding, perch hopping, preening, and drinking), novel object exploration and colour learning task between LD and dLAN. Non-parametric Mann-Whitney’s U test compared the changes in fat scores (0 to 5) between LD and dLAN. Two-way repeated measures analysis of variance (2-way RM ANOVA) followed by Bonferroni post hoc test, if ANOVA tested a significant difference assessed the effect of the light condition (factor 1: LD vs. dLAN) and the retention interval (factor 2: 1 h vs. 24 h) on reward search time. Pearson and Spearman correlation coefficient was calculated to show correlation between two variables (colour learning and sleep). We tested 24 h rhythmicity in the daily behaviors by unimodal non-linear cosinor regression analysis (y = A + [B.cos (2π (x − C)/24)]), where A, B, and C denote the mesor (mean 24 h behavioral values), amplitude (maximum change in behavior levels relative to the mesor) and acrophase (the estimated time of peak of behavior) of 24 h rhythm, respectively (Prabhat et al., 2019) [20]. The significance of the cosinor regression was calculated by using the number of samples, R2 values and numbers of predictors (https://www.danielsoper.com/statcalc/calculator.aspx?id=15; Soper 2013, 29 August 2021). An extra sum of squares F-test also determined significant differences in the rhythm waveform parameters (i.e., mesor, amplitude and acrophase) of daily behaviors between light conditions, as reported in several publications [13,20]. The alpha was set at 0.05 for statistical significance.
3. Results
3.1. Effect on Daily Behaviors
We assessed the effect of light at night on daily feeding, drinking, preening, and perch-hopping behavior in male zebra finches to look at the direct effect of illuminated nights. LD birds restricted all of their activity in light period only, with dLAN birds extended their behaviors in the night time also. Almost 30–50% of total behavioral activities were found to be in the nighttime in dLAN birds (Table 1). As expected from earlier experiments, all behaviors showed robust significant daily rhythm under LD controls (Cosinor regression: p < 0.0001; Figure 2). We found persistence in rhythmicity in all other behaviors under dLAN as well (Cosinor regression: p < 0.0001, p < 0.01; Figure 2). However, the feeding duration was arrythmic under dLAN (Cosinor regression: p = 0.065). Interestingly, between LD and dLAN the rhythm waveforms were significantly different. Specifically, amplitude of feeding and preening frequency and mesor of feeding and drinking frequency was significantly lower in dLAN compared to LD (F-test: p < 0.0001, p < 0.05; Figure 2). We did not find any difference in the acrophase in behaviors between both the light conditions (F-test: p > 0.05; Figure 2).
3.2. Effect on Body Mass, Fat Score, Food Intake and Crop Seed Content
We measured changes in body mass, fat score, and food intake to assess the effects of light at night on both feeding behavior and body condition. There was significant increase in the body mass of dLAN birds (15–20%) as compared to LD birds after six weeks (Student’s t-test: p < 0.05; Table 1). This might be due to increase in fat deposition hence overall fat score under light at night condition (Mann-Whitney U test: p < 0.05; Table 1). The range of fat score under LD was 0–1 and dLAN 2–5. Interestingly, dLAN birds ate more food at night (Video S4) and the crop seed content was also higher at the lights on as compared to LD (Student’s t-test: p < 0.05; Table 1).
3.3. Effect on Sleep and Novel Object Exploration
We assayed various sleep parameters and novel object exploration to look at the effect of light at night on mood. Sleep was restricted to night time only in both the groups however, due to presence of light during nighttime birds under dLAN showed various activity at night unlike LD. Specifically, dLAN birds showed significant delay in sleep latency (Student’s t-test: t14 = 4.18, p = 0.0009) and advanced awakening latency (Student’s t-test: t14 = 4.17, p = 0.0009). dLAN birds significantly increased number of sleep bouts, which also suggest more disturbance in sleep behavior or fragmentation of sleep as compared to LD (Student’s t-test: t14 = 8.55, p < 0.0001), hence total sleep duration was also less in dLAN (Student’s t-test: t14 = 9.74, p < 0.0001). To look at the effect on mood, we performed the novel object exploration test which has already been established to assay mood in rodents and other songbirds [8,13,14,34]. There was significant effect of light at night on object exploration as dLAN birds did not associate (spent time) with novel object (Student’s t-test: p < 0.05). Specifically, only one out of eight dLAN birds reached a score of 4 (perch closely with novel object) compared to all the birds under LD (Mann-Whitney U test: p < 0.05; Figure 3). The total perch hops during NOE were also limited to LD birds as most of the dLAN birds did not hop during test period (Student’s t-test: p < 0.05).
3.4. Colour Learning Task
A total of six and five out of eight birds in LD and dLAN, respectively, performed the colour learning task under LD and dLAN, respectively. Birds under dLAN took significantly fewer trials to learn the rewarded colour (7 to 14) than LD (2 to 5) (Mann-Whitney U test: p = 0.004; Figure 4). There was significant effect of light condition (F(1, 9) = 10.91, p = 0.009) on search time during memory test but not of retention interval (F(1, 9) = 0.36, p = 0.56) and light condition x retention interval interaction (2-way RM ANOVA: F(1, 9) = 2.79, p = 0.13). Specifically, dLAN birds took more time to perform the correct task in 24 h retention interval memory test than LD birds (Bonferroni posttest: p < 0.01). Table 2 summarizes the colour learning and memory task of Zebra Finches in both the light conditions. No incorrect response (error) was observed under LD in 1-h RI as compared to 5 times error in dLAN in completed trials. Interestingly, out of 50 trials (25 in 1-h RI and 24-h RI each) under dLAN, 10 times birds choose not to attempt the memory task which shows the unresponsiveness or lack of motivation under dLAN. Also, there was a significant negative correlation of total sleep duration with number of trials (Spearman’s correlation: r2 = 0.60; p = 0.0054, Figure 4) and search time in 24 h retention memory test irrespective of the light condition (Pearson’s correlation: r2 = 0.87; p < 0.00001, Figure 4).
4. Discussion
One of the key issues of unnatural LAN is the ability to enable activities that allow animals including humans to eat, forage, and sleep throughout the day that can lead to circadian disruption and altered health conditions [6]. Here, we show that a dim light at night (dLAN) for six weeks has negative effects on sleep behavior, physiology, daily behavioral rhythms, novel object exploration, and colour learning task on diurnal adult male zebra finches. We used dLAN at ecological relevant levels (measured from different areas of university campus at nighttime to correlate with the emerging illuminated night environment. We found that dLAN extended the daily activity into the night which affected the overall rhythmicity of putative behaviors [17,18]. Constant bright light conditions have shown to affect circadian rhythmicity in activity and singing behavior in songbirds [15,20,35,36]. Specifically, the feeding behavior became arrhythmic, as birds ate almost similar amounts in the night time compared to the day time (Table 1). Studies on mammals and songbirds have shown that ‘wrong time’ of eating can lead to fattening and increase in body mass via bright and dim light at night conditions in mammals and songbirds [37,38,39,40]. Similar to these findings, zebra finches under dLAN increased their body mass and fat score without changing the total food intake. Light at night induced fattening and lipid accumulation in the liver was observed in female zebra finches [17,27]. This could be the result of change in rhythmicity and allocation of energy expenditure throughout the day (Figure 2). Artificial light at night have shown to affect the foraging behavior in urban areas in feral pigeons as they extend the feeding and activity behavior after sunset [41]. The presence of light at night was the only variable changed between LD and dLAN groups, and effects on activity and feeding behavior and clock gene expression has been observed under constant light and dim light at night conditions in zebra finches [17,21]. However, we cannot discount the effects of other factors as we have not carried out any molecular analysis.
The persistence of activities into nighttime resulted in reduced total sleep duration and fragmentation of sleep behavior in zebra finches under illuminated night conditions, similar to reduced sleep in Indian house crows, female zebra finches, European blackbirds and great tits under dim light at night [9,14,16,18,24,42]. Interestingly, birds were more disturbed due to the presence of night illumination, hence more numbers of sleep bouts (i.e., more sleep-awake situations were observed). The presence of light at dusk and dawn time also affected the sleep and awakening latency, as dLAN birds slept later and woke up nearly an hour earlier than LD. Overall, we can suggest that a dim light at night disturbed the sleep-wake rhythm and resulted in decreased sleep behavior. We did not measure the serum melatonin and oxalic acid in the present study. However, a decline in night melatonin and oxalic acid level has been reported in songbirds under dLAN [9,14,17,18]. Male zebra finches with decreased sleep behavior also showed reduced novel object exploratory behavior. Birds showed almost no involvement and did not spent time with novel object under dLAN, similar to the loss of interest and neophobia such as behavior under light at night condition (constant bright light and dim light) in diurnal Grass Rats and songbirds [8,13,14,34]. Neophobia can be helpful in the novel environment in which a guarded response to the novel object or environment will be better for survival. However, in a laboratory environment with lack of predators, an exploratory response to novel object can be linked with interest, mood, and higher brain functions [13,14].
A similar result was observed in motivation dependent learning and memory tasks. There was a concurrent decline in the colour learning and memory performance in zebra finches under dLAN, as evidenced by lesser participation, more error, and longer search duration, specifically during memory retention interval and non-responsiveness to perform tasks (Figure 4, Table 2). Since birds ate at the nighttime throughout the experiment under dLAN as examined by crop seed content (Table 1), we argue the decrease in motivation under dLAN in the given reward-based task. It could be the cumulative effect of light at night as it impacts the learning and memory in Indian House Crows [10,14]. Also, birds performed better during 24 h RI of colour learning under LD condition similar to Indian House Crows in spatial learning and memory tasks [10]. Interesting findings from the study revealed that, the memory retention decreased as search time duration increased during 24 h RI in dLAN birds compared to 1 h RI. One of the reasons could be that light can also affect the various brain regions including thalamus, midbrain, hippocampus and pallium, hence the effect of light at night on the decline in cognitive performance including memory tasks cannot be discounted [43,44]. A decline in cognitive performance in spatial and pattern learning task was correlated with the activity of learning- and memory-associated brain regions by the expression of ZENK (a neuronal activity marker) in the hippocampus and pallium under LL and reduced neurogenesis via BDNF and DCX protein expression under dLAN in Indian House Crows [10,14]. Illuminated nights regulate the higher brain functions by attenuating the dopamine secretion pathway as shown in mammals and songbirds [10,45]. Similar to earlier studies, we argue that negative effects on colour learning and memory might be due to the changes in the brain area involved rather than direct effect of excess lighting at night [46].
Overall, our results are similar to effects on female Zebra Finches in Batra et al., 2019 [17]. The female birds showed extreme fattening after three weeks of dLAN as compared to male birds which took five to six weeks to get fattened in the present study. Studies on great tits have shown that female tits spent a greater proportion of night awake in response to artificial light at night environment [16]. There are few limitations of the present study which could have attributed to the presence or absence of effects of dim light at night on behavior. First, a good number of sample size would have given a better understanding of learning and memory task as 5 out of 16 birds did not participate in the colour learning task. Second, the effects of different spectrum and intensity of light at night has not been addressed via present study as we did not measure the spectral properties of light. Therefore, we cannot discount the effects caused by specific wavelength of dim light at night (24), rather than white light. Third, a very dim light at night can also influence behavior and physiology in birds, hence unable to differentiate between no light and very dim light might not have provided a better explanation for the changes in behavior. We propose that future studies should focus on delineating the molecular mechanism through which light at night affects behavior and physiology in animals. The present study tried to address the effect of overly lit environment at wrong time on learning and memory which might be an important issue for almost all vertebrates in these days of rapid urbanization. Similar studies on learning and memory would increase the focus on higher brain function. For these purposes, zebra finches would be an excellent model, as it is diurnal, breeds in laboratory condition and many studies have used zebra finches to understand behaviouoral biology, reproductive biology, and physiology [28].
5. Conclusions
We show that dLAN negatively affects zebra finches, a diurnal species, at the level of daily behavioral rhythms, sleep behavior, physiology, object exploration and learning and memory tasks. We suggest that alteration in rhythm and decreased sleep behavior affected overall mood and motivation to interact with novel objects and increased non-performance in learning and memory tasks. Insights from these results may be useful in understanding the impact of nighttime illumination, such as in an overly lit urban habitat, on daily behavior, physiology, and advanced brain functions in animals.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/birds2040028/s1. Video S1: Novel object exploration; Video S2: Error in colour learning and memory task; Video S3: Correct response in colour learning and memory task; Video S4: Zebra Finch feeding at night under dLAN.
Author Contributions
Conceptualization, V.K., S.K.B. and A.P.; methodology, S.K.B., A.P.; validation, A.P., M.K., A.K. and S.K.B.; formal analysis, S.K.B. and A.P.; investigation, A.P., M.K. and A.K.; resources, S.K.B.; data curation, A.P., M.K. and A.K.; writing—original draft preparation, A.P., M.K. and A.K.; writing—review and editing, S.K.B. and A.P.; supervision, V.K. and S.K.B.; project administration, S.K.B.; funding acquisition, SERB (Science and Engineering Research Board, New Delhi, India). All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by SERB (Science and Engineering Research Board, New Delhi, India), grant number SERB- CRG/2019/000321 to S.K.B. A.P. received a research associate fellowship from the SERB, New Delhi.
Institutional Review Board Statement
This project and study was approved by Institutional Animal Ethical Committee of Chaudhary Charan Singh University (IAEC-2020/3; 25-11-2020).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data will be available with the corresponding author upon request.
Acknowledgments
We acknowledge the generous funding from SERB and help of Nidhi Yadav in the experiment. We are grateful to all three reviewers for helpful comments and suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kumar, V.; Singh, B.P.; Rani, S. The bird clock: A complex, multi-oscillatory and highly diversified system. Biol. Rhythm Res. 2004, 35, 121–144. [Google Scholar] [CrossRef]
- Dunlap, J.C.; Loros, J.J.; DeCoursey, P.J. Chronobiology: Biological Timekeeping, 1st ed.; Sinauer Associates: Sunderland, MA, USA, 2004. [Google Scholar]
- Albrecht, U. Circadian clocks and mood-related behaviors. Handb. Exp. Pharmacol. 2013, 217, 227–239. [Google Scholar]
- Golombek, D.A.; Rosenstein, R.E. Physiology of circadian entrainment. Physiol. Rev. 2010, 90, 1063–1102. [Google Scholar] [CrossRef] [Green Version]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Ann. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunn, R.M.; Blask, D.E.; Coogan, A.N.; Figueiro, M.G.; Gorman, M.R.; Hall, J.E.; Hansen, J.; Nelson, R.J.; Panda, S.; Smolensky, M.H.; et al. Health consequences of electric lighting practices in the modern world: A report on the National Toxicology Program’s workshop on shift work at night, artificial light at night, and circadian disruption. Sci. Total Environ. 2017, 607, 1073–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonken, L.K.; Workman, J.L.; Walton, J.C.; Weil, Z.M.; Morris, J.S.; Haim, A.; Nelson, R.J. Light at night increases body mass by shifting the time of food intake. Proc. Nat. Acad. Sci. USA 2010, 107, 18664–18669. [Google Scholar] [CrossRef] [Green Version]
- Fonken, L.K.; Aubrecht, T.G.; Meléndez-Fernández, O.H.; Weil, Z.M.; Nelson, R.J. Dim light at night disrupts molecular circadian rhythms and increases body weight. J. Biol. Rhythms 2013, 28, 262–271. [Google Scholar] [CrossRef]
- De Jong, M.; Ouyang, J.Q.; van Grunsven, R.H.; Visser, M.E.; Spoelstra, K. Do wild great tits avoid exposure to light at night? PLoS ONE 2016, 11, e0157357. [Google Scholar] [CrossRef]
- Taufique, S.T.; Jha, N.A.; Kumar, V. Circadian rhythm determines the timing of activity, and ingestive and grooming behaviours in Indian house crows, Corvus splendens. Curr. Sci. 2016, 110, 897–901. [Google Scholar]
- Bedrosian, T.A.; Nelson, R.J. Influence of the modern light environment on mood. Mol. Psychiatr. 2013, 18, 751–757. [Google Scholar] [CrossRef]
- Raap, T.; Casasole, G.; Costantini, D.; AbdElgawad, H.; Asard, H.; Pinxten, R.; Eens, M. Artificial light at night affects body mass but not oxidative status in free-living nestling songbirds: An experimental study. Sci. Rep. 2016, 6, 35626. [Google Scholar] [CrossRef]
- Jha, N.A.; Kumar, V. Effect of no-night light environment on behaviour, learning performance and personality in zebra finches. Anim. Behav. 2017, 132, 29–47. [Google Scholar] [CrossRef]
- Taufique, S.T.; Prabhat, A.; Kumar, V. Illuminated night alters hippocampal gene expressions and induces depressive-like responses in diurnal corvids. Eur. J. Neurosci. 2018, 48, 3005–3018. [Google Scholar] [CrossRef]
- Singh, J.; Rani, S.; Kumar, V. Functional similarity in relation to the external environment between circadian behavioral and melatonin rhythms in the subtropical Indian weaver bird. Horm. Behav. 2012, 61, 527–534. [Google Scholar] [CrossRef]
- Raap, T.; Pinxten, R.; Eens, M. Light pollution disrupts sleep in free-living animals. Sci. Rep. 2015, 5, 13557. [Google Scholar] [CrossRef] [Green Version]
- Batra, T.; Malik, I.; Kumar, V. Illuminated night alters behaviour and negatively affects physiology and metabolism in diurnal zebra finches. Environ. Poll. 2019, 254, 112916. [Google Scholar] [CrossRef]
- Batra, T.; Malik, I.; Prabhat, A.; Bhardwaj, S.K.; Kumar, V. Sleep in unnatural times: Illuminated night negatively affects sleep and associated hypothalamic gene expressions in diurnal zebra finches. Proceed. R. Soc. B 2020, 287, 20192952. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Harpole, C.E.; Trivedi, A.K.; Cassone, V.M. Circadian regulation of bird song, call, and locomotor behavior by pineal melatonin in the zebra finch. J. Biol. Rhythms 2012, 27, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Prabhat, A.; Jha, N.A.; Taufique, S.T.; Kumar, V. Dissociation of circadian activity and singing behavior from gene expression rhythms in the hypothalamus, song control nuclei and cerebellum in diurnal zebra finches. Chronobiol. Int. 2019, 36, 1268–1284. [Google Scholar] [CrossRef]
- Prabhat, A.; Malik, I.; Jha, N.A.; Bhardwaj, S.K.; Kumar, V. Developmental effects of constant light on circadian behaviour and gene expressions in zebra finches: Insights into mechanisms of metabolic adaptation to aperiodic environment in diurnal animals. J. Photochem. Photobiol. B 2020, 211, 111995. [Google Scholar] [CrossRef]
- Kumar, J.; Malik, S.; Bhardwaj, S.K.; Rani, S. Bright light at night alters the perception of daylength in Indian weaver bird (Ploceus philippinus). J. Exp. Zool. A Ecol. Integr. Physiol. 2018, 329, 488–496. [Google Scholar] [CrossRef]
- Navara, K.J.; Nelson, R.J. The dark side of light at night: Physiological, epidemiological, and ecological consequences. J. Pineal Res. 2007, 43, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.Q.; de Jong, M.; van Grunsven, R.H.; Matson, K.D.; Haussmann, M.F.; Meerlo, P.; Visser, M.E.; Spoelstra, K. Restless roosts: Light pollution affects behavior, sleep, and physiology in a free-living songbird. Glob. Chang. Biol. 2017, 23, 4987–4994. [Google Scholar] [CrossRef]
- Crusio, W.E. Genetic dissection of mouse exploratory behaviour. Behav. Brain Res. 2001, 125, 127–132. [Google Scholar] [CrossRef]
- Mishra, I.; Kumar, V. The quantity–quality trade-off: Differential effects of daily food availability times on reproductive performance and offspring quality in diurnal zebra finches. J. Exp. Biol. 2019, 222, jeb196667. [Google Scholar] [CrossRef] [Green Version]
- Prabhat, A.; Batra, T.; Kumar, V. Effects of timed food availability on reproduction and metabolism in zebra finches: Molecular insights into homeostatic adaptation to food-restriction in diurnal vertebrates. Horm. Behav. 2020, 125, 104820. [Google Scholar] [CrossRef]
- Griffith, S.C.; Ton, R.; Hurley, L.L.; McDiarmid, C.S.; Pacheco-Fuentes, H. The ecology of the Zebra finch makes it a great laboratory model but an outlier amongst passerine birds. Birds 2021, 2, 60–76. [Google Scholar] [CrossRef]
- Yadav, A.; Kumar, R.; Tiwari, J.; Kumar, V.; Rani, S. Sleep in birds: Lying on the continuum of activity and rest. Biol. Rhythm Res. 2017, 48, 805–814. [Google Scholar] [CrossRef]
- Raap, T.; Pinxten, R.; Eens, M. Artificial light at night causes an unexpected increase in oxalate in developing male songbirds. Conserv. Physiol. 2018, 6, coy005. [Google Scholar] [CrossRef] [PubMed]
- Steinmeyer, C.; Schielzeth, H.; Mueller, J.C.; Kempenaers, B. Variation in sleep behaviour in free-living blue tits, Cyanistes caeruleus: Effects of sex, age and environment. Anim. Behav. 2010, 80, 853–864. [Google Scholar] [CrossRef]
- Mainwaring, M.C.; Beal, J.L.; Hartley, I.R. Zebra finches are bolder in an asocial, rather than social, context. Behav. Process. 2011, 87, 171–175. [Google Scholar] [CrossRef]
- Boogert, N.J.; Giraldeau, L.A.; Lefebvre, L. Song complexity correlates with learning ability in zebra finch males. Anim. Behav. 2008, 76, 1735–1741. [Google Scholar] [CrossRef]
- Fonken, L.K.; Kitsmiller, E.; Smale, L.; Nelson, R.J. Dim nighttime light impairs cognition and provokes depressive-like responses in a diurnal rodent. J. Biol. Rhythms 2012, 27, 319–327. [Google Scholar] [CrossRef]
- Cassone, V.M.; Westneat, D.F. The bird of time: Cognition and the avian biological clock. Front. Mol. Neurosci. 2012, 5, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, N.; Mishra, I.; Komal, R.; Rani, S.; Kumar, V. Circannual testis and moult cycles persist under photoperiods that disrupt circadian activity and clock gene cycles in spotted munia. J. Exp. Biol. 2017, 220, 4162–4168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonken, L.K.; Nelson, R.J. The effects of light at night on circadian clocks and metabolism. Endocr. Rev. 2014, 35, 648–670. [Google Scholar] [CrossRef]
- Plano, S.A.; Casiraghi, L.P.; García Moro, P.; Paladino, N.; Golombek, D.A.; Chiesa, J.J. Circadian and metabolic effects of light: Implications in weight homeostasis and health. Front. Neurol. 2017, 8, 558. [Google Scholar] [CrossRef] [Green Version]
- Coomans, C.P.; van den Berg, S.A.; Houben, T.; van Klinken, J.B.; van den Berg, R.; Pronk, A.C.; Havekes, L.M.; Romijn, J.A.; van Dijk, K.W.; Biermasz, N.R.; et al. Detrimental effects of constant light exposure and high-fat diet on circadian energy metabolism and insulin sensitivity. FASEB J. 2017, 27, 1721–1732. [Google Scholar] [CrossRef] [Green Version]
- Borniger, J.C.; Weil, Z.M.; Zhang, N.; Nelson, R.J. Dim light at night does not disrupt timing or quality of sleep in mice. Chronobiol. Int. 2013, 30, 1016–1023. [Google Scholar] [CrossRef]
- Leveau, L.M. Artificial light at night (ALAN) is the main driver of nocturnal feral pigeon (Columba livia f. domestica) foraging in urban areas. Animals 2020, 10, 554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, A.; Samplonius, J.M.; Schlicht, E.; Valcu, M.; Kempenaers, B. Artificial night lighting rather than traffic noise affects the daily timing of dawn and dusk singing in common European songbirds. Behav. Ecol. 2014, 25, 1037–1047. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V. Avian photoreceptors and their role in the regulation of daily and seasonal physiology. Gen. Comp. Endocrinol. 2015, 220, 13–22. [Google Scholar] [CrossRef]
- Hattar, S.; Kumar, M.; Park, A.; Tong, P.; Tung, J.; Yau, K.W.; Berson, D.M. Central projections of melanopsin-expressing retinal ganglion cells in the mouse. J. Comp. Neurol. 2006, 497, 326–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romeo, S.; Viaggi, C.; Di Camillo, D.; Willis, A.W.; Lozzi, L.; Rocchi, C.; Capannolo, M.; Aloisi, G.; Vaglini, F.; Maccarone, R.; et al. Bright light exposure reduces TH-positive dopamine neurons: Implications of light pollution in Parkinson’s disease epidemiology. Sci. Rep. 2013, 3, 1395. [Google Scholar] [CrossRef] [PubMed]
- Taufique, S.T.; Kumar, V. Differential activation and tyrosine hydroxylase distribution in the hippocampal, pallial and midbrain brain regions in response to cognitive performance in Indian house crows exposed to abrupt light environment. Behav. Brain Res. 2016, 314, 21–29. [Google Scholar] [CrossRef] [PubMed]
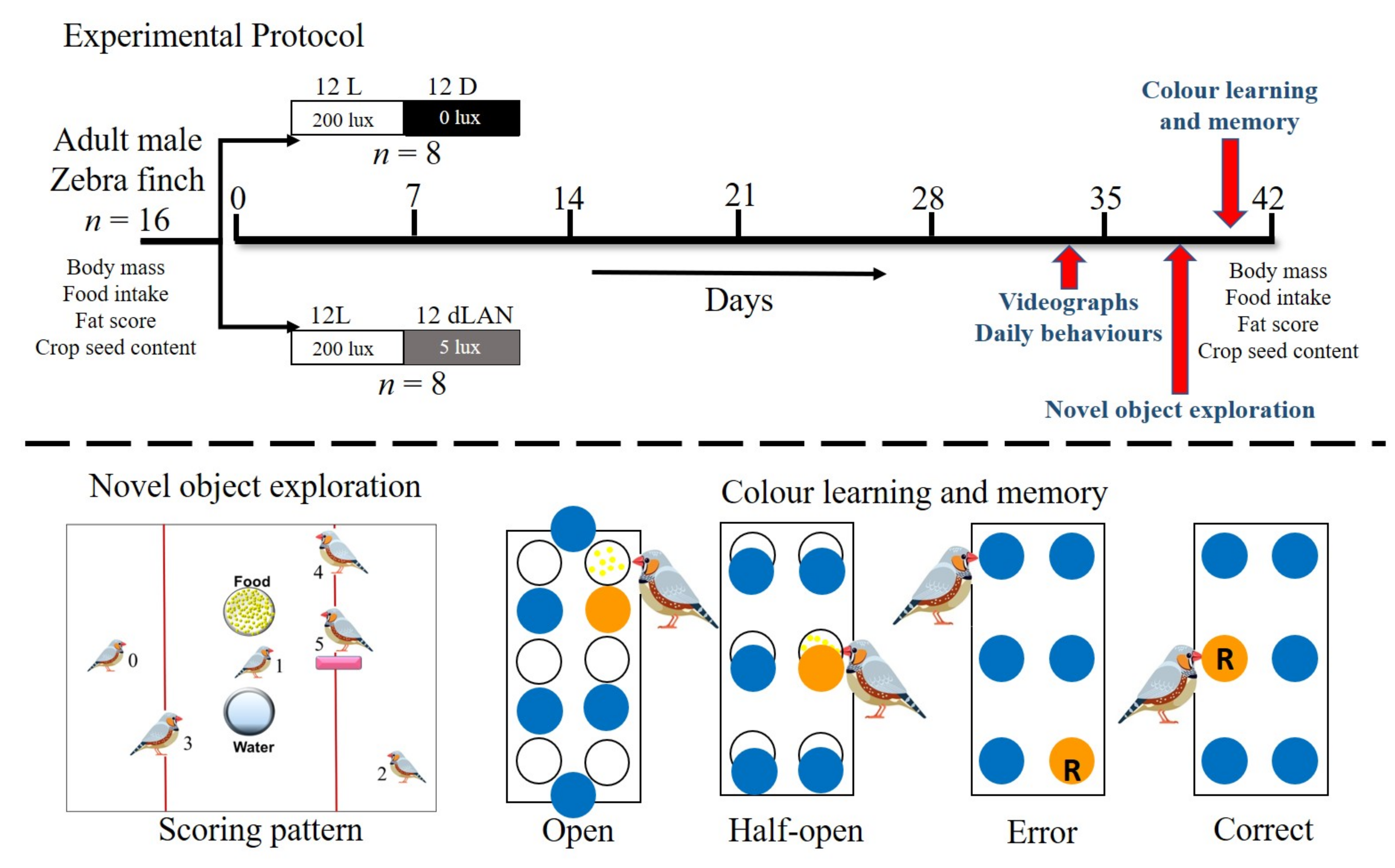
Figure 1.
Experimental protocol: Adult male zebra finches, Taeniopygia guttata (n = 16) were subjected to dim light at night (L: 200 ± 10 lux, dLAN = 5 ± 1 lux) or darkness at night (L: 200 ± 10 lux, D < 0.1 lux) for six weeks at 22 ± 2 °C with food and water ad libitum. We recorded physiological parameters (body mass, food intake and fat score) and crop seed content at the start and end of the experiment. We videographed daily behaviors of birds at the end of week five. Birds were tested for novel object exploration and colour learning and memory task at the end of week six (for details see materials and methods section).
Figure 1.
Experimental protocol: Adult male zebra finches, Taeniopygia guttata (n = 16) were subjected to dim light at night (L: 200 ± 10 lux, dLAN = 5 ± 1 lux) or darkness at night (L: 200 ± 10 lux, D < 0.1 lux) for six weeks at 22 ± 2 °C with food and water ad libitum. We recorded physiological parameters (body mass, food intake and fat score) and crop seed content at the start and end of the experiment. We videographed daily behaviors of birds at the end of week five. Birds were tested for novel object exploration and colour learning and memory task at the end of week six (for details see materials and methods section).

Figure 2.
A non-linear regression rhythm plot of feeding, perch-hopping, drinking and preening behavior of adult male zebra finches (Taeniopygia guttata) under 12 h light: 12 h darkness (12L:12D) with absolute dark nights ((LD: L = 200 lux; D < 0.1 lux), and 12 h light: 12 h dim light at night (dLAN: L = 200 lux; D = 5 lux). The white bar represents light period and black and grey bar represents darkness (D) and dim light at night (dLAN) period, respectively. The solid and dotted line represents rhythmicity and arrhythmicity in behavior, respectively as determined by cosinor regression analysis. The waveform characteristics are defined as mesor (Mes), amplitude (Amp) and acrophase (Acr) of significant 24-h rhythmic behaviors. The F-test determined the difference between LD and dLAN parameters. The * symbol shows significance tested by Student’s t-test. For significance, alpha was set at 0.05.
Figure 2.
A non-linear regression rhythm plot of feeding, perch-hopping, drinking and preening behavior of adult male zebra finches (Taeniopygia guttata) under 12 h light: 12 h darkness (12L:12D) with absolute dark nights ((LD: L = 200 lux; D < 0.1 lux), and 12 h light: 12 h dim light at night (dLAN: L = 200 lux; D = 5 lux). The white bar represents light period and black and grey bar represents darkness (D) and dim light at night (dLAN) period, respectively. The solid and dotted line represents rhythmicity and arrhythmicity in behavior, respectively as determined by cosinor regression analysis. The waveform characteristics are defined as mesor (Mes), amplitude (Amp) and acrophase (Acr) of significant 24-h rhythmic behaviors. The F-test determined the difference between LD and dLAN parameters. The * symbol shows significance tested by Student’s t-test. For significance, alpha was set at 0.05.

Figure 3.
Sleep and novel object exploration behavior in adult male zebra finches (Taeniopygia guttata) under 12 h light: 12 h darkness (12L:12D) and 12 h light: 12 h dim light at night (12L:12dLAN) as described in Figure 1. The upper panel shows changes in the four different sleep parameters (sleep latency, awakening latency, numbers of sleep bout and total sleep duration) and photomicrographs showing front and back sleep. Lower panel shows the results of NOE test and scoring pattern based on bird’s position with respect to the novel object. Box-whisker plot represents the range of highest to lowest values. Black circle represents individual value. The * symbol shows significance tested by Student’s t-test, set at p < 0.05 (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001).
Figure 3.
Sleep and novel object exploration behavior in adult male zebra finches (Taeniopygia guttata) under 12 h light: 12 h darkness (12L:12D) and 12 h light: 12 h dim light at night (12L:12dLAN) as described in Figure 1. The upper panel shows changes in the four different sleep parameters (sleep latency, awakening latency, numbers of sleep bout and total sleep duration) and photomicrographs showing front and back sleep. Lower panel shows the results of NOE test and scoring pattern based on bird’s position with respect to the novel object. Box-whisker plot represents the range of highest to lowest values. Black circle represents individual value. The * symbol shows significance tested by Student’s t-test, set at p < 0.05 (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001).

Figure 4.
The colour learning and memory performance of adult male zebra finches (Taeniopygia guttata) under 12 h light: 12 h darkness (12L:12D), and 12 h light: 12 h dim light at night (12L:12dLAN) as described in Figure 1. We analyzed number of trials to learn the task and search time during 1-h and 24-h memory retention test under LD and dLAN represented in box-whisker plot. Black circle represents individual value. Relationship between number of trials and search time with total sleep duration. The * symbol shows significance tested by Student’s t-test (number of trials), 2-way RM-ANOVA (search time) and correlation between colour learning and sleep. Alpha was set at 0.05 (** p < 0.01, **** p < 0.0001).
Figure 4.
The colour learning and memory performance of adult male zebra finches (Taeniopygia guttata) under 12 h light: 12 h darkness (12L:12D), and 12 h light: 12 h dim light at night (12L:12dLAN) as described in Figure 1. We analyzed number of trials to learn the task and search time during 1-h and 24-h memory retention test under LD and dLAN represented in box-whisker plot. Black circle represents individual value. Relationship between number of trials and search time with total sleep duration. The * symbol shows significance tested by Student’s t-test (number of trials), 2-way RM-ANOVA (search time) and correlation between colour learning and sleep. Alpha was set at 0.05 (** p < 0.01, **** p < 0.0001).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparison of body physiology and daily behaviors in male zebra finches (Taeniopygia guttata) under Light: Dark, LD (12L:12D; L = 200 ± 10 lux, D < 0.1 lux) and dim light at night, dLAN (12L:12dLAN; L = 200 ± 10 lux, dLAN = 5 ± 1 lux) light condition. Values are presented in mean ± SEM unless specified. Parametric Student’s t test and non-parametric Mann-Whitney test compared the light conditions. Bold and unbold values represent presence and absence of significance. Alpha was set at 0.05 for significance.
Table 1.
Comparison of body physiology and daily behaviors in male zebra finches (Taeniopygia guttata) under Light: Dark, LD (12L:12D; L = 200 ± 10 lux, D < 0.1 lux) and dim light at night, dLAN (12L:12dLAN; L = 200 ± 10 lux, dLAN = 5 ± 1 lux) light condition. Values are presented in mean ± SEM unless specified. Parametric Student’s t test and non-parametric Mann-Whitney test compared the light conditions. Bold and unbold values represent presence and absence of significance. Alpha was set at 0.05 for significance.
| A. Effect of dLAN on change in body mass, food intake, crop seed content, and fat score (final-initial value) | ||||
| Parameter | LD | dLAN | Analysis | |
| ∆Body mass (g) | −0.08 ± 0.27 | 1.25 ± 0.35 | t14 = 2.98; p = 0.01 | |
| ∆Food intake (g) | Day: −0.24 ± 0.31 Night: 0.0 ± 0.0 | Day: −1.2 ± 0.29 Night: 1.02 ± 0.21 | t14 = 3.62; p = 0.003 t14 = 4.71; p < 0.001 | |
| ∆Crop seed content (at lights on) | 0 | 12.88 ± 4.24 | t14 = 3.03; p = 0.009 | |
| ∆Fat score (median) | 0 | 3 | Mann-Whitney U = 10.50; p = 0.02 | |
| B. Effect of dLAN on daily activity (perch-hopping and preening), feeding, drinking, and preening | ||||
| Parameter | Time | LD | dLAN | Analysis |
| Feeding frequency | Day | 26.06 ± 3.58 | 20.88 ± 3.56 | t14 = 1.03; p = 0.32 |
| Night | 0.0 ± 0.0 | 14.06 ± 1.99 | t14 = 7.06; p < 0.0001 | |
| Feeding duration (min) | Day | 105.9 ± 19.48 | 70.91 ± 15.42 | t14 = 1.40; p = 0.18 |
| Night | 0.0 ± 0.0 | 39.07 ± 4.97 | t14 = 7.86; p < 0.0001 | |
| Perch hopping | Day | 1752 ± 405.5 | 1728 ± 552.8 | t13 = 0.03; p = 0.97 |
| Night | 0.0 ± 0.0 | 401.9 ± 174.6 | t13 = 2.30; p = 0.04 | |
| Drinking frequency | Day | 20.44 ± 1.32 | 26.19 ± 6.07 | t14 = 0.93; p = 0.37 |
| Night | 0.0 ± 0.0 | 8.88 ± 2.62 | t14 = 3.39; p = 0.004 | |
| Drinking duration (min) | Day | 8.68 ± 2.35 | 8.84 ± 3.27 | t14 = 0.04; p = 0.97 |
| Night | 0.0 ± 0.0 | 3.66 ± 1.63 | t14 = 2.24; p = 0.04 | |
| Preening frequency | Day | 38.13 ± 7.26 | 30.81 ± 5.41 | t14 = 0.43; p = 0.43 |
| Night | 0.0 ± 0.0 | 10.25 ± 1.69 | t14 = 6.06; p < 0.0001 | |
Table 2.
The colour learning and memory task of male zebra finches (Taeniopygia guttata) under LD (12L:12D) and dLAN (12L:12dLAN) light condition as described in Figure 1. The table shows colour association memory performance after 1-h RI (retention interval) and 24-h RI (retention interval). In total, 5 trials of 10 min were given during memory retention. ✓ = correct response; X = no. of incorrect response in particular trial; N = not attempted in particular trials.
Table 2.
The colour learning and memory task of male zebra finches (Taeniopygia guttata) under LD (12L:12D) and dLAN (12L:12dLAN) light condition as described in Figure 1. The table shows colour association memory performance after 1-h RI (retention interval) and 24-h RI (retention interval). In total, 5 trials of 10 min were given during memory retention. ✓ = correct response; X = no. of incorrect response in particular trial; N = not attempted in particular trials.
| LD | dLAN | |||
|---|---|---|---|---|
| Bird | 1-h RI | 24-h RI | 1-h RI | 24-h RI |
| 1 | ✓ ✓ ✓ ✓ ✓ | ✓ X ✓ ✓ ✓ | ----- | ----- |
| 2 | ----- | ----- | ✓ ✓ N ✓ N | ✓ ✓ ✓ N N |
| 3 | ✓ ✓ ✓ ✓ ✓ | ✓ ✓ ✓ ✓ ✓ | ----- | ----- |
| 4 | ✓ ✓ ✓ ✓ ✓ | ✓ ✓ ✓ ✓ ✓ | ✓ ✓ N N ✓ | ✓ ✓ N ✓ N |
| 5 | ✓ ✓ ✓ ✓ ✓ | ✓ ✓ ✓ ✓ ✓ | ✓ X X ✓✓ | X ✓ X ✓ N |
| 6 | ----- | ----- | ✓ ✓ ✓ ✓ ✓ | ✓ ✓ ✓ ✓ ✓ |
| 7 | ✓ ✓ ✓ ✓ ✓ | ✓ ✓ ✓ ✓ ✓ | ----- | ----- |
| 8 | ✓ ✓ ✓ ✓ N | X ✓ ✓ ✓ ✓ | ✓ ✓ N ✓ X | ✓ ✓ ✓ X ✓ |
| X | X | |||
| X | X | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Prabhat, A.; Kumar, M.; Kumar, A.; Kumar, V.; Bhardwaj, S.K. Effects of Night Illumination on Behavior, Body Mass and Learning in Male Zebra Finches. Birds 2021, 2, 381-394. https://doi.org/10.3390/birds2040028
AMA Style
Prabhat A, Kumar M, Kumar A, Kumar V, Bhardwaj SK. Effects of Night Illumination on Behavior, Body Mass and Learning in Male Zebra Finches. Birds. 2021; 2(4):381-394. https://doi.org/10.3390/birds2040028
Chicago/Turabian StylePrabhat, Abhilash, Mayank Kumar, Ashwani Kumar, Vinod Kumar, and Sanjay K. Bhardwaj. 2021. "Effects of Night Illumination on Behavior, Body Mass and Learning in Male Zebra Finches" Birds 2, no. 4: 381-394. https://doi.org/10.3390/birds2040028